




Transition Period Management and Nutrition
Most of the metabolic diseases of dairy cows - milk fever, ketosis, retained placenta (RP), and displacement of the abomasum - occur within the first 2 wk of lactation. This feature was taken from the proceedings of the High Plains Dairy Conference. A full link to the proceedings is provided below. 12 April 2009
12 April 2009
 32 minute read
32 minute read
Introduction
The etiology of many of those metabolic diseases that are not clinically apparent during the first 2 wk of lactation, such as laminitis, can be traced back to insults that occurred in early lactation. In addition to metabolic diseases, the overwhelming majority of infectious diseases, in particular mastitis, become clinically apparent during the first 2 wk of lactation.
Three basic physiologic functions must be maintained during the periparturient period if disease is to be avoided. These are:
- A strong immune system;
- Normocalcemia; and
- Feed intake during the days before and after calving.
Both metabolic disease and infectious disease incidence are greatly increased whenever one or more of these physiological functions is impaired. Each of these 3 physiological factors influence the risk for development of the most common diseases encountered during the periparturient period. A cow failing in any 1 of these 3 areas is also more likely to suffer a failure in another vital function. Because other speakers at this conference will address management and dietary issues affecting maintenance of feed intake in peripartueint cows (cow comfort, grouping, social factors, high straw vs. higher starch, monensin, number of days in the close-up group), this will not be discussed in this review.
Maintaining a Strong Immune System Around the Time of Calving
Does Milk Production Affect Immune Status?
Neutrophil and lymphocyte function is diminished in the periparturient period, especially in the dairy cow (Kehrli et al. 1989 a,b; Detilleux et al., 1994). The onset of milk production imposes tremendous challenges to the mechanisms responsible for energy, protein, and mineral homeostasis in the cow. Negative energy, protein, and/or mineral balance and hormonal fluxes associated with the onset of lactation may be responsible for the immunosuppression observed in periparturient dairy cattle.
Mastectomy of pregnant dairy cows removes the impact of milk production while maintaining endocrine and other changes associated with late pregnancy and parturition. Mastectomy would be expected to improve immune function in the periparturient dairy cow, if milk production is an immunosuppressive factor. Using 10 mastectomized and 8 intact multiparous Jersey cows (all intact cows developed milk fever) we assessed the ability of neutrophils to kill microbes as assessed by neutrophil myeloperoxidase activity during the periparturient period (Kimura et. al., 1999). Neutrophil myeloperoxidase activity decreased equally before parturition in both groups. While there was a quick recovery of neutrophil myeloperoxidase activity in mastectomized cows, there was no recovery in intact cows after parturition throughout the study, which lasted until d 20 postpartum. Lymphocyte production of gamma-interferon in vitro declined significantly at parturition in intact cows but did not decrease significantly in the mastectomized cows. In intact cows, all T cell subset populations (i.e. CD3, CD4, CD8, and gamma-delta positive cells) decreased as a percentage of total peripheral blood mononuclear cells (PBMC) at the time of parturition, while the percentage of monocytes increased. These population changes have previously been shown to be associated with the immune suppression commonly observed in periparturient cows. Mastectomy eliminated these changes in leukocyte subsets (Kimura et al., 2002b).
These results suggest: 1) the mammary gland may produce substances which directly affect immune cell populations, or 2) metabolic demands associated with the onset of lactation negatively impact the composition and functioning of circulating PBMC populations. Two metabolic factors were greatly impacted by mastectomy. Mastectomy eliminated hypocalcemia at parturition. Plasma non-esterified fatty acid (NEFA) concentration rose dramatically in intact cows at calving and did not return to baseline levels for > 10 d. In contrast NEFA concentration in mastectomized cow plasma rose only slightly at calving and returned to baseline levels the day after calving. It is clear that the intact cow mobilizes a much larger amount of body fat than does the mastectomized cow, suggestive of a severe negative energy balance at the onset of lactation.
Hypocalcemia and Mastitis Susceptibility
Milk fever cows are at increased risk of developing mastitis (Curtis et al., 1983). Why? Calcium (Ca) is necessary for proper contraction of muscle. Severe hypocalcemia prevents skeletal muscle contraction to the point that the clinical syndrome known as milk fever occurs. Muscle contraction is reduced by any decrease in blood Ca; however it must be severe before we observe the downer cow. Daniel et al. (1983) demonstrated that contraction rate and strength of the smooth muscle of the intestinal tract is directly proportional to blood Ca concentration. Hypocalcemia reduces abomasal contraction, which causes the abomasum to fill with gas and become displaced. Of course lack of effective fiber is the other main contributor to displacement of the abomasum. A major reason for the current trend toward high straw close-up diets is the ability of these types of diets to prevent displacement of the abomasum.
It should be kept in mind that the sphincter at the end of the teat is also a muscle. The teat sphincter smooth muscle must contract if closure of the teat end is to occur. If low blood Ca reduces teat sphincter contraction, the teat canal may remain open, inviting environmental pathogens to enter the mammary gland. While we generally associate milk fever with the day of calving, we have demonstrated that many cows remain subclinically hypocalcemic for the first week of lactation (Goff et al., 1996). Hypocalcemic cows tend to spend more time lying down than do normocalcemic animals. Again, this could increase teat end exposure to environmental opportunists.
Hypocalcemia also acts as a stressor to the cow. Cows typically exhibit a 3-4 fold increase in plasma cortisol as part of the act of initiation of parturition. However subclinically hypocalcemic cows may have 5-7 fold increases in plasma cortisol on the day of calving and the typical milk fever cow may exhibit plasma cortisol concentrations that are 10 –15 fold higher than pre-calving plasma cortisol concentration (Horst and Jorgensen, 1982). Cortisol is generally considered a powerful immune suppressive agent and likely exacerbates the immune suppression normally observed in the periparturient period. Exacerbates, rather than causes, because most studies suggest that immune suppression begins 1-2 wk before calving (Kehrli et al., 1989 a,b; Ishikawa et al., 1987; Kashiwazaki et al., 1985), and the cortisol surge is fairly tightly confined to the day of calving and perhaps the day after calving.
Recently, we have found a direct association between hypocalcemia and failure of immune cells to become activated when they encounter a stimulus such as bacteria (Kimura et al., 2006). When an immune cell such as a lymphocyte encounters a bacterial antigen at its surface it triggers the release of Ca from organelles within the cell. This begins the process by which the lymphocyte will produce antibodies, bactericidal peptides, etc., to kill the bacteria. A rise in intracellular Ca is a key early feature in immune cell activation. Peripheral blood mononuclear cells were obtained from multiparous dairy cows spanning a period of 2 wk before and 2 wk after parturition. The cells were exposed to activating stimuli in vitro and intracellular Ca release from intracellular stores was measured. The intracellular Ca released in response to the activation signal declined as Ca demand for lactation became more intense and recovered as plasma Ca normalized. Hypocalcemia, which is common in periparturient dairy cows, was causing decreased intracellular Ca stores in PBMC. Our data suggest this caused a blunted intracellular Ca release response to an immune cell activation signal and helps explain how hypocalcemia contributes to periparturient immune suppression.
Ketosis and Mastitis Susceptibility
Ketosis is diagnosed whenever there are elevated levels of ketones in the blood, urine, or milk of a cow. The disease is always characterized by a decline in blood glucose as well. In lactation, the amount of energy required for maintenance of body tissues and milk production exceeds the amount of energy the cow can obtain from her diet; especially in early lactation when dry matter intake is still low.
The fresh cow is also in negative protein balance shortly after calving. Generally this is not perceived to be as big a problem as the negative energy balance of early lactation, but the typical cow will lose 37 lb of body protein during the first 2 wk of lactation. Much of this body protein is being used to support the amino acid and glucose requirements of milk production (Paquay et al., 1972). Therefore, in many respects, the dairy cow in early lactation is in a physiological state comparable to that of humans and rodents with prolonged protein-calorie restriction. Glutamine is the most abundant free amino acid in human muscle and plasma and is utilized at high rates by rapidly dividing cells, including leucocytes, to provide energy and optimal conditions for nucleotide biosynthesis.
A few studies have been done on lymphocytes isolated directly from normal cows and cows with clinical and subclinical ketosis and then placed into culture. Leukocytes of cows with clinical symptoms and the highest concentration of ketones and free fatty acids in blood responded with the lowest levels of interferons alpha and gamma to 3 interferon inducers: Newcastle Disease virus, phytohemagglutinin, and concanavalin A (Kandefer-Szerszen et al., 1992).
Zerbe et al. (2000) examined the relationship between liver triacyl glycerol (TAG) content and functional properties of neutrophils of dairy cows in the peripartum period. Increased liver TAG content, > 40 mg/g, which was considered the upper level of normal, went in parallel with a reduced expression of function-associated surface molecules on blood neutrophils. Moreover, in cows with high liver TAG levels the antibody-independent and -dependent cellular cytotoxicity of blood polymorphonucleotites (PMN) was markedly reduced. PMN also were less capable of reactive oxygen species generation after stimulation with phorbol myristate acetate.
Bottom line – the cow with ketosis has almost no resistance to infectious disease.
What is the energy cost to mount an immune response?
Little to no work has been done to examine this issue in cattle. However, in humans suffering from severe infection causing sepsis (various degrees of fever, increased white blood cell (WBC) count, and acute phase protein production), the resting energy expenditure (determined by indirect calorimetry), increased progressively over the first week of the infection to around 40 per cent above normal and was still elevated 3 wk from the onset of illness. As an aside; over a 3-wk period patients lost 13 per cent of their total body protein (Plank and Hill, 2000).
No such measurements have been reported for cattle. However if we extrapolate and speculate, we can go through a few calculations. Maintenance energy for a 600 kg dairy cow is approximately 9.7 mcal net energy/d. If the cow must also increase energy expenditure 40 per cent to mount an inflammatory response as in humans, the energy requirement increases by nearly 4 mcal/d. This is roughly equivalent to a requirement that the cow consume an additional 2.4 kg of diet (assuming a lactating diet that provided 1.65 mcal NEl/kg). Can the periparturient cow, already in negative energy balance, be expected to successfully mount a rapid immune response? If she is in fact in negative protein balance as well, will her immune system produce the immunoglobulins and acute phase proteins necessary to fight an infection while it is still in the acute phase to prevent it from escalating to a clinical infection?
Feed Intake, Immune Function, and Susceptibility to Retained Placenta, Metritis, and Endometritis
There has been some association between the development of RP and the incidence of mastitis (Emanuelson et al., 1993; Peeler et al., 1994). We have recently confirmed studies begun by Gunnink (1984) that suggest the two are likely linked, because both are due to immune suppression in affected cows. Gunnink’s theory suggested the fetal placenta must be recognized as foreign tissue and rejected by the immune system after parturition to cause expulsion of the placenta. We hypothesized that impaired neutrophil function causes RP. We examined the ability of neutrophils to recognize fetal cotyledon tissue as assessed by a chemotaxis assay, which utilized a placental homogenate obtained from a spontaneously expelled placenta as the chemoattractant. Neutrophil killing ability was also estimated by determining myeloperoxidase activity in isolated neutrophils. Blood samples were obtained from 142 periparturient dairy cattle in 2 herds. Twenty cattle developed RP (14.1 per cent).
Neutrophils isolated from blood of cows with RP had significantly lower neutrophil function in both assays prior to calving and this impaired function lasted for 1-2 wk after parturition. Addition of antibody directed against interleukin-8 (IL-8) to the cotyledon preparation used as a chemoattractant inhibited chemotaxis by 41 per cent, suggesting one of the chemoattractants present in the cotyledon at parturition is IL-8. At calving plasma IL-8 concentration was lower in RP cows (51 ± 12 pg/ml) than in cows expelling the placenta normally (134 ± 11 pg/ml; Kimura et al., 2002a). These data suggest neutrophil function determines whether or not the cow will develop RP. These data also suggest that depressed production of IL-8 may be a factor affecting neutrophil function in cows developing RP. This suppressed immune system could also explain why the same cows are more susceptible to mastitis. Retained placenta probably does not cause mastitis, but is symptomatic of a depressed immune system.
At calving the cow’s reproductive tract is exposed to bacteria, even in the cleanest of environments. The cow survives because her WBC provide protection from infection. Neutrophils provide the first line of defense, moving out of the blood whenever and wherever bacteria invade body tissue. Once in the infected tissue, the neutrophils ingest the bacteria and release enzymes and free radical compounds onto the bacteria to kill them. Occasionally, the neutrophils don’t succeed in killing the intruder. The immune system then calls on macrophages and lymphocytes, which work together to produce antibodies and other anti-bacterial factors. Production of these factors takes a little more time, but will eventually eliminate most infections the neutrophils can’t handle.
In immune-compromised cows the bacteria are not kept in check and grow to large numbers in the uterus, causing a condition known as metritis. Around 20-30 per cent of cows develop metritis, which is characterized by a foul-smelling, red-brown, watery discharge from the uterus within 10-14 d after calving. It is often, but not always, accompanied by a fever. Hammon et al. (2006) demonstrated that neutrophils of cows with metritis are significantly less able to kill bacteria (measured by a neutrophil iodination assay) than neutrophils from cows without metritis. The surprise was that poor neutrophil function was evident in these cows the day of calving – before lactation began and before any bacteria could have entered the uterus.
Endometritis is a uterine problem characterized by inflammation of the lining of the uterus lasting more than 3-4 wk after calving. Studies suggest 40-50 per cent of cows can have endometritis at 4 wk after calving. These cows are less likely to successfully breed back. An interesting study by Kim et al. (2005) compared the peripartum immune responses of dairy cows that developed endometritis by 4 wk postpartum (n = 11) to cows that did not develop this disease (n = 19). Blood samples were collected 1 wk before calving, just after or during calving, and then at wk 1, 2, 3, and 4 postpartum. The leukocytes from cows that developed endometritis were significantly less phagocytic than those from control cows at all sampling time points (P < 0.01). In the study of Hammon et al. (2006), cows with endometritis and sub-clinical endometritis had poorer neutrophil function than cows with a healthy uterus - from the time they calved until the diagnosis of endometritis was made 4 wk later. Cows in this study developing metritis, or diagnosed with subclinical or clinical endometritis, had higher NEFA levels in their blood than did cows with a healthy uterus. Surprisingly, the NEFA levels were significantly higher for at least 2 wk prior to calving, suggesting these cows were mobilizing body fat even before calving. Individual cow feed intake data revealed those cows that were going to develop uterine health problems ate significantly less feed than cows that would maintain a healthy uterus. This difference in feed intake existed at least 1 wk prior to calving. Urton et al. (2005) were able to demonstrate that feeding behavior prior to calving was a major risk factor for metritis. In this study, for every 10 min/d reduction in time spent eating in the weeks prior to calving, the risk of developing metritis doubled. Though uterine diseases are diagnosed in early lactation, it now appears that the health of the uterus is greatly influenced by feed intake and avoidance of negative energy at the end of the dry period.
Micronutrients and the Immune System
The biggest culprit affecting the immune system are the hormone changes associated with calving and the negative energy and protein balances of cows in early lactation. The energy and protein imbalances are not easily fixed. Deficiencies of any of the micronutrients will result in immune function loss as well. To that end the following guidelines are utilized by me to prevent the short term deficiencies in vitamins and minerals we sometimes see in dairy cattle.
Unless you are using a pasture-based system, feed 4000 IU vitamin E/d for the 2-3 wk before calving and 2000 IU vitamin E the first 2 wk of lactation. This recommendation is based on the work of Weiss et al. (1997) and our own observations on plasma vitamin E levels at calving (Goff and Stabel, 1990). In the study of Weiss the addition of vitamin E greatly decreased the incidence of mastitis. If these data are only half correct, the vitamin E is a cheap investment.
Feed 0.3 mg/kg selenium, the legal limit. Add trace minerals in adequate amounts. Farms should get in the routine habit of assessing the adequacy of their trace mineral program once or twice each year by harvesting liver from a cow that has died rapidly due to trauma, lightning, etc., or is going to slaughter; and then having the liver sample analyzed for trace mineral content. Liver is the best indicator of the status of copper, zinc, and manganese supplementation. Blood is generally unreliable for these minerals. Whole blood selenium is a good indicator of selenium deficiency. In some cases inorganic trace minerals do not seem capable of meeting the mineral needs of animals. This is often due to the presence of substances in the diet that interfere with absorption of the inorganic trace minerals. Organic sources of trace minerals are less susceptible to antagonists and should be considered when lab tests suggest deficiency in the face of good levels of inorganic minerals. Another strategy is to insure adequate trace mineral status by routine use of the more expensive organic trace minerals, in place of expensive lab tests. Iron can be antagonistic to trace mineral absorption. It is also a strong pro-oxidant; this increases the need for vitamin E. Dietary iron greater than 800 mg/kg diet and water iron greater than 0.7-2 mg/kg is too much. We think this causes oxidative stress in animals and reduces immune cell function.
Maintaining Normal Blood Calcium Concentration
Inadequate blood Ca concentrations can cause a cow to lose the ability to rise to her feet, as Ca is necessary for nerve and muscle function. The result is the metabolic disease known as milk fever; though it is more properly termed periparturient hypocalcemia or periparturient paresis, as an elevated body temperature is not typically observed. Surveys in the USA suggest around 5 per cent of cows will develop milk fever each year and the incidence of subclinical hypocalcemia – blood Ca values between 8 and 5.5 mg/dl during the perparturient period - is around 50 per cent in older cows. Milk fever and subclinical milk fever should be considered gateway diseases that greatly reduce the chance for full productivity in the ensuing lactation. Hypocalcemia reduces rumen and abomasal motility, increasing the risk of abomasal displacement. Hypocalcemia reduces feed intake so that greater body fat mobilization occurs in early lactation. Hypocalcemia reduces all muscle contractions including the teat sphincter muscle responsible for closure of the teat orifice after milking; thus increasing the risk of mastitis. More recently we have demonstrated hypocalcemia directly impairs immune cell response to an activating stimulus.
Ca Homeostasis and Monitoring for Hypocalcemia
Blood Ca in the adult cow is maintained between 8.5 – 10 mg/dl (2.1 – 2.5 mmol/L). Typically the nadir in blood Ca concentration occurs between 12 and 24 hr after calving and blood samples obtained around this time can reveal the extent of hypocalcemia experienced by a dairy herd. Nearly 25 per cent of heifers will have blood Ca concentration below 8 mg/dl. About 50 per cent of older cows will fall into this category. In well-managed herds following a good anionic salt program, or other effective milk fever control measures, the above figures can be cut in half and the number of cows exhibiting clinical milk fever can be reduced to 1per cent or less. Acute hypocalcemia can also occur under many conditions involving infections, such as mastitis or metritis, especially if endotoxins are elaborated. As a rule the blood Ca concentration is below 8, but above 6 mg/dl. It is due to re-distribution of Ca within organs and will not be discussed further, other than to be a reminder that not all hypocalcemic cows have the syndrome known as milk fever.
In order to prevent blood Ca from decreasing at the onset of lactation, the cow must replace extracellular Ca lost to milk. She does this by withdrawing Ca from bone and by increasing the efficiency of absorption of dietary Ca. The dairy cow (as are most mammals) is programmed to go into a state of lactational osteoporosis, mobilizing bone Ca to help her achieve normocalcemia in early lactation. This will typically result in loss of 9-13 per cent of her skeletal Ca in the first month of lactation (which is reversible in later lactation). Though it might stress her bones, the main objective - to maintain normocalcemia - can be achieved. Bone Ca mobilization is regulated by parathyroid hormone (PTH), which is produced whenever there is a decline in blood Ca. Renal tubular reabsorption of Ca is also enhanced by PTH. However the total amount of Ca that can be recovered by reducing urinary Ca excretion is relatively small, as only small amounts of Ca are typically lost to urine each day. A second hormone, 1,25-dihydroxyvitamin D, is required to stimulate the intestine to efficiently absorb dietary Ca. This hormone is made from vitamin D by the kidney –but only in response to an increase in blood PTH. Put simply, hypocalcemia and milk fever occur when cattle do not extract enough Ca from their bones and diet to replace the Ca lost to milk. Several nutritional factors are involved in the breakdown of Ca homeostasis that results in milk fever.
Factors Impairing Calcium Homeostasis at the Cellular Level
Metabolic Alkalosis
Metabolic alkalosis predisposes cows to milk fever and subclinical hypocalcemia (Craige and Stoll, 1947). Metabolic alkalosis blunts the response of the cow to PTH (Gaynor et al., 1989; Goff et al., 1991; Phillippo et al., 1994). We now believe the conformation of the PTH receptor is altered during metabolic alkalosis, rendering the tissues less sensitive to PTH. Lack of PTH responsiveness by bone tissue prevents effective utilization of bone canaliculi fluid Ca, sometimes referred to as osteocytic osteolysis, and prevents activation of osteoclastic bone resorption. Failure of the kidneys to respond to PTH also reduces renal reabsorption of Ca from the glomerular filtrate. More importantly, the kidneys fail to convert 25-hydroxyvitamin D to 1,25-dihydroxyvitamin D. Therefore enhanced intestinal absorption of dietary Ca, that normally would help restore blood Ca to normal, fails to be instituted. Metabolic alkalosis is largely the result of a diet that supplies more cations (potassium (K), sodium (Na), Ca, and magnesium (Mg)) than anions (chloride (Cl), sulfate (SO4), and phosphate (PO4)) to the blood. In simplest terms, a disparity in electrical charge in body fluids occurs in animals fed these diets, because a greater number of positively charged cations enter the blood than negatively charged anions. To restore electroneutrality to this high cation, positively charged blood, a positive charge in the form of a hydrogen ion (H+) must be lost from the blood compartment. A reduction in H+ concentration is equivalent to an increase in the pH of the blood (Stewart, 1983). For a more detailed description of how dietary cation-anion balance influences blood pH, the reader is referred to recent reviews on this subject (Constable, 1999; Goff, 2000). Adding readily absorbable anions to the diet increases the total negative charges in the blood, allowing more H+ to exist and the blood pH decreases – it is more acidic.
Hypomagnesemia
Cow plasma Mg concentration is normally between 1.8 and 2.4 mg/dl (0.75 and 1.0 mmol/L). Hypomagnesemia affects Ca metabolism in 2 ways: 1. by reducing PTH secretion in response to hypocalcemia (Littledike et al., 1983) and 2. by reducing tissue sensitivity to PTH (Rude, 1998).
The integrity of the interaction between PTH and its receptor is vital to Ca homeostasis. Hypomagnesemia, independent of metabolic alkalosis, can also interfere with the ability of PTH to act on its target tissues. When PTH binds its receptor on bone or kidney tissues, it normally initiates activation of adenylate cyclase, resulting in production of the second messenger, cyclic AMP. PTH-receptor interactions should also cause activation of phospholipase C in some tissues, resulting in production of the second messengers diacylglycerol and inositol 1,4,5-triphosphate. Both adenylate cyclase and phospholipase C have a Mg++ binding site which must be occupied by a Mg ion for full activity (Rude, 1998). In man, it is well recognized that hypomagnesemia can cause hypocalcemia and that Mg therapy alone restores the serum Ca concentration to normal; Ca and/or vitamin D therapy are ineffective (Rude, 1998). Field evidence suggests that blood Mg concentrations below 0.65 mmol/L in the periparturient cow will increase the susceptibility of cows to hypocalcemia and milk fever (van de Braak et al., 1987).
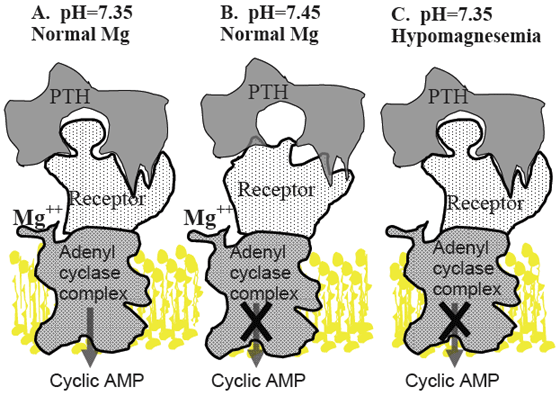
Panel A. Under normal conditions, PTH released in response to hypocalcemia interacts with its receptor, located on the surface of bone and kidney cells, in a lock and key fashion. This stimulates G-proteins and adenylate cyclase (adenylate cyclase complex) resulting in production of cyclic AMP, which acts as a second messenger within the cytosol of target cells. This initiates mechanisms such as bone Ca resorption and renal production of 1,25-dihydroxyvitamin D to restore blood Ca concentration to normal levels.
Panel B. Alkalotic conditions induced by high potassium diets induce a change in the shape of the PTH receptor protein so that it is less able to recognize and bind PTH, resulting in failure to activate the cell by producing cyclic AMP.
Panel C. Mg is required for function of the adenylate cyclase complex. Hypomagnesemia reduces ability of PTH stimulated cells to produce cylic AMP, resulting in failure to activate the cell.
Maintenance of normal plasma Mg concentration is nearly totally dependent on a constant influx of Mg from the diet. Magnesium is well absorbed from the small intestine of young calves and lambs. As the rumen and reticulum develop these sites become the main, and perhaps the only, sites for net Mg absorption (Martens and Rayssiguier, 1980). Magnesium absorption from the rumen is dependent on the concentration of Mg in solution in the rumen fluid and the integrity of the Mg transport mechanism (Martens and Gabel, 1986).
The soluble concentration of Mg in rumen fluid is obviously dependent on the Mg content of the diet. However, Mg solubility declines sharply as rumen pH rises above 6.5 and solubility can be a problem on higher forage diets. Forages also can contain trans-aconitic acid. A metabolite of trans-aconitic acid, tricarballylate, can complex Mg, is resistant to rumen degradation, and may play a role in hypomagnesemic tetany (Cook et al., 1994).
Active transport of Mg across the rumen wall is necessary when diet Mg is not in great supply. Unfortunately, high K concentration in the rumen fluid depolarizes the apical membrane of the rumen epithelium reducing the electromotive potential needed to drive Mg across the rumen wall (Martens and Schweigel, 2000). Thus a ration that might otherwise be adequate in Mg results in a Mg deficient state when diet K is excessive.
A second pathway for absorption of Mg exists that is not affected by K. Unfortunately, this passive transport process only operates at high rumen fluid Mg concentrations, which allow Mg to flow down a concentration gradient into the extracellular fluids of the cow (Martens and Schweigel, 2000). The concentration of Mg in rumen fluid needed to utilize concentration gradient driven absorption of Mg is greater than 4 mmol/L (Care et al., 1984; Ram et al., 1998). The minimum level of Mg required in the diet to prevent negative Mg balance in the face of high K levels in ruminants is approximately 0.35 per cent (Ram et al., 1998). Thus, Mg content of the close-up dry cow ration and the early lactation ration should be between 0.35 and 0.4 per cent, as insurance against the possibility that the active transport processes for Mg absorption are impaired.
Assessing Mg Status
Sampling the blood of several cows within 12 hr after calving provides an effective index of Mg status of the periparturient cows. If serum Mg concentration is not at least 1.8 mg/dl (0.8 mmol/L) it suggests inadequate dietary Mg absorption and that hypomagnesemia may be limiting productivity as well as contributing to hypocalcemia in the herd. Cows with blood Mg between 1.15 and 1.8 mg/dl (0.5 and 0.8 mmol/L) have few obvious clinical symptoms, though they often are slow to eat and are not producing milk up to their potential. Clinical signs of hypomagnesemia, such as recumbency, convulsions, nystagmus; are only observed when blood Mg falls below 0.4-0.5 mmol/L. Tetany is generally accompanied by severe hypocalcemia. Hypomagnesemia is very amenable to prevention by increasing dietary Mg content and form.
Reducing Dietary Cation-anion Difference to Prevent Hypocalcemia
In theory all the cations and anions in a diet are capable of exerting an influence on the electrical charge of the blood. The major cations present in feeds and the charge they carry are Na (+1), K (+1), Ca (+2), and Mg (+2). The major anions and their charges found in feeds are Cl (-1), SO4 (-2), and phosphate (assumed to be -3). Cations or anions present in the diet will only alter the electrical charge of the blood if they are absorbed into the blood. Trace elements present in diets are absorbed in such small amounts that they are of negligible consequence to acid-base status. Organic acids, such as the volatile fatty acids, are generally absorbed in the undissociated form so that they carry both a positive and negative charge into the blood. They also are rapidly metabolized within the liver, so they have only a small effect on general acid-base balance under most circumstances.
The difference between the number of cation and anion particles absorbed from the diet determines the general acid-base balance of the body and therefore, the pH of the blood. The cation-anion difference of a diet is commonly described in terms of mEq/kg DM (some authors prefer to use mEq/100 g diet DM) of just Na, K, Cl, and SO4; although it must be kept in mind that Ca, Mg, and P absorbed from the diet will also influence blood pH. Experimental evidence from our laboratory (Goff et al., 2004) and the meta-analysis of Charbonneau et al. (2006) support the concept that dietary SO4 is only about 60 per cent as effective as Cl as an acidifying agent in the diet of the prepartum cow, suggesting an appropriate DCAD equation could be (Na + K) – (Cl + 0.6 S). While DCAD equations provide a theoretical basis for dietary manipulation of acid-base status they are not necessary for formulation of mineral content of prepartum dairy cow rations in this author’s opinion because, with the exception of K and Cl, the rate of inclusion of the other macrominerals can be set at fixed rates- as outlined below.
The USA National Research Council (NRC, 2000) requirement for Na in the diet of a late gestation cow is about 0.12 per cent. A small amount of salt is added to the diet to prevent pica, which often is manifest as a desire to drink urine from the floor. Unlimited access to NaCl is to be avoided in late gestation because it will increase the risk of udder edema, not because it greatly affects acid-base status.
At least 2 studies have clearly demonstrated that inclusion of Ca in the diet at NRC required levels, or several fold above NRC required levels, does not influence the degree of hypocalcemia experienced by the cow at calving (Goff and Horst, 1997; Beede et al., 2001). It appears from these studies that the close-up diet Ca concentration should be maintained between 0.85 and 1.0 per cent Ca.
To ensure adequate concentrations of Mg in the blood of the periparturient cow the dietary Mg concentration should be 0.35-0.4 per cent. This higher dietary Mg concentration allows the cow to take advantage of passive absorption of Mg across the rumen wall. Be sure the Mg source you are using is available – many MgO sources I have encountered in the field are not.
Dietary P concentration should be fed at a level to meet the NRC requirement for P in the late gestation cow. This is generally about 0.4 per cent P for most cows. A diet supplying more than 80 g P/d (Barton, 1978; Kichura et al., 1982) will block renal production of 1,25-dihydroxyvitamin D and will actually cause milk fever.
Dietary S must be kept above 0.22 per cent to ensure adequate substrate for rumen microbial amino acid synthesis. Corn (maize) silage diets are notoriously low in S. Diet S should be kept below 0.4 per cent to avoid possible neurological problems associated with S toxicity (Gould et al., 1991).
Now, with the exception of K and Cl, the variables in the various proposed DCAD equations have become more or less fixed. The key to milk fever prevention (at least with Holstein cows) is to keep K as close to the NRC requirement of the dry cow as possible (about 1.0 per cent diet K). The key to reduction of subclinical hypocalcemia, not just milk fever, is to add Cl to the ration to counteract the effects of even low diet K on blood alkalinity. For formulation purposes, the concentration of Cl required in the diet to acidify the cow is approximately 0.5 per cent less than the concentration of K in the diet. In other words, if diet K can be reduced to 1.3 per cent, the Cl concentration of the diet should be increased to 0.8 per cent. If dietary K can only be reduced to 2.0 per cent the diet Cl would need to be roughly 1.5 per cent to acidify the cow. This level of Cl in the diet is likely to cause a decrease in dry matter intake. Chloride sources differ in their palatability. Since achieving low dietary K can be difficult, it is prudent to use a palatable source of Cl when formulating the diet. Ammonium chloride (or ammonium sulfate) can be particularly unpalatable when included in rations with a high pH. At higher pH, a portion of the ammonium cation is converted to ammonia, which is highly irritating when smelled by the cow. Prilling the Cl (and SO4) salts reduces the unpleasant taste of the salts. In our experience hydrochloric acid has proved the most palatable source of anions. Hydrochloric acid can be extremely dangerous to handle when it is procured as a liquid concentrate. Several North American companies now manufacture hydrochloric acid based anion supplements, which are safe to handle. Some also include Mg in a highly soluble form to reduce the possibility that hypomagnesemia is present in the herd and contributing to hypocalcemia.
These are simply guidelines for anion supplementation used by this author that are based on inclusion of Ca, Na, S, Mg, and P at the levels outlined above. Urine pH of the cows provides a cheap and fairly accurate assessment of blood pH and can be a good gauge of the appropriate level of anion supplementation (Jardon, 1995). Urine pH on high cation diets is generally above 8.2. Limiting dietary cations will reduce urine pH only a small amount (down to ~7.8). For optimal control of subclinical hypocalcemia, the average pH of the urine of Holstein cows should be between 6.2 and 6.8; which essentially requires addition of anions to the ration. In Jersey cows the average urine pH of the close-up cows has to be reduced to between 5.8 and 6.3 for effective control of hypocalcemia. If the average urine pH is between 5.0 and 5.5, excessive anions have induced an uncompensated metabolic acidosis and the cows will suffer a decline in dry matter intake. Urine pH can be checked 48 hr or more after a ration change. Urine samples should be free of feces and made on midstream collections to avoid alkalinity from vaginal secretions. Anion supplemented diets are generally fed for the last 3 wk before calving; though the length of time these diets need to be fed to induce a compensated metabolic acidosis is no more than 4-5 d.
How do I interpret urine pH? Let’s say you check urine pH of ten close-up cows. You are interested in the average – there is always one odd ball cow. Don’t adjust the diet to accommodate the one cow!!
- Scenario 1 - average pH = 6.3 + 0.6 – Congratulations you have induced a compensated metabolic acidosis.
- Scenario 2 - average pH = 7.4 + 0.5 – Add anion source in 0.25 lb/cow increments, wait 3 d and check pH.
- Scenario 3 - average pH 5.2 + 0.5 – You have induced an uncompensated metabolic acidosis = sick cows! Reduce anion source by 0.5 lb/cow and check urine pH in 3 d.
- Scenario 4 - 4 cows at pH 5.2 and 6 cows at pH 7.8
– Cows at pH 5.2 today ate well this morning but will not this afternoon, due to uncompensated metabolic acidosis. The cows at pH 7.8 went off feed yesterday from over-acidification, but will likely eat today and be over-acidifed tomorrow. Reduce anion source by 0.5 lb/cow, wait 4-5 d, check pH, and start increasing anion source by 0.25 lb increments as needed.
Further Reading
| - | You can view the full conference proceedings by clicking here. |
April 2009


