






Feeding Rumininally-Protected Choline to Transition Dairy Cows
Choline, also known as trimethyl ethanolamine, is an essential component of mammalian diets as it is required for normal function of cells, says this University of Florida IFAS Publication. 14 June 2009
14 June 2009
 15 minute read
15 minute read
![]()
Introduction
It was discovered by Andreas Strecker in 1862, but it remained unrecognized as an essential nutrient for humans until 1998, when dietary reference intakes were established. Choline was initially determined to be essential in the prevention of fatty liver in rats and dogs (Best and Huntsman, 1932). Choline is key compound for the synthesis of 2 important molecules, phosphatidylcholine and acetylcholine (Figure 1).
Methylation Pathways
(DNA, proteins, lipids)
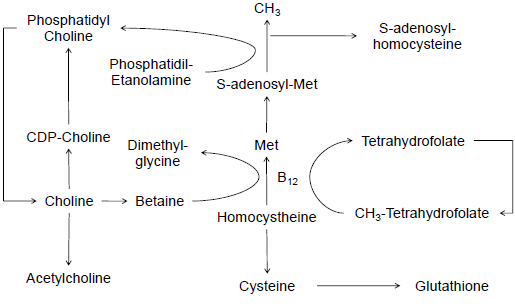
Figure 1. Metabolic pathways for choline and its relationships with folic acid and methionine as CH3 carrier for methylation pathways
Phosphatidylcholine is the main phospholipid in ruminants and it is critical for lipid absorption and transport, cell membrane structure, cell signaling, and synthesis of lipoproteins (Zeisel and Holmes-McNary, 2001). The second compound synthesized from choline, acetylcholine, is a neurotransmitter in the central and peripheral nervous system and critical in the neuromuscular junctions for muscle contractions. Acetylcholine is synthesized from choline and acetyl-coA in neurons. Because choline is a critical component for synthesis of phospholipids and neurotransmitters, it supports structural integrity and signaling roles in cell membranes, influences cholinergic neurotransmission and cognition, and it is a major source for methyl-groups via betaine for methylation pathways (Figure 1).
However, in ruminant nutrition, choline is typically viewed as a nonessential component, despite indications that only up to 30 per cent of the estimated required choline is provided by duodenal flow of choline (Erdman, 1992). Therefore, most of the choline required by dairy cows must be produced by de novo synthesis through the sequential methylation of phosphatidyletanolamine, with the methyl groups being supplied by S-adenosyl-L-methionine (Zeisel and Holmes-McNary, 2001). Nevertheless, because de novo synthesis can generate “sufficient” methyl donors for synthesis of phosphatidylcholine, the presence of this pathway does not mean that increased dietary supply and intestinal absorption of choline is not required for optimum cow health and metabolism.
Ruminal metabolism
Choline that reaches the small intestine of cows is in the form of phosphatidylcholine present in rumen microbes, primarily protozoa (Broad and Dawson, 1976). High concentrate diets typically reduce protozoa numbers, which could limit the supply of choline to cows. Sharma and Erdman (1989a) estimated the in vitro ruminal degradation of choline present in feedstuffs typically fed to cows.
The mean estimates of rumen degradable choline were 79.4, 84.7, 82.9, 83.8, 98.0, and 98.6 per cent for barley, cottonseed meal, fish meal, soybean meal, choline stearate, and choline chloride, respectively. The authors suggested that dietary choline fed in unprotected forms such as choline-rich feedstuffs or choline chloride would only marginally increase the postruminal flow of choline in dairy cows. In fact, the same group conducted 3 experiments to quantify the extension of ruminal degradation of choline and the subsequent duodenal flow in dairy cows (Atkins et al., 1988). In the first two experiments, feeding different concentrations of dietary choline (range from 0 to 5 g/kg of diet DM) had no effect on yields of milk or fat-corrected milk, but supplemental choline tended to increase fat yield and percentage.
In experiment 3, ruminally and duodenally canulated steers were dosed with 27 g/d supplemental choline and duodenal flow of choline was measured. Steers consumed on average 13.1 kg of DM/d, which resulted in a choline intake of 46.7 g/d in the unsupplemented group. Dosing steer in the rumen with 27 g of choline increased intake to 72.6 g/d. Although duodenal flow was altered, it increased by only 3 g/d (13.7 vs. 16.6 g/d) with adding 27 g of choline in the rumen. The calculated apparent rumen degradability of choline tended to be greater for the animals that received 27 g/d of choline (77.1 vs. 70.6 per cent) than the unsupplemented animals.
When lactating dairy cows received 0, 10 and 20 g of supplemental choline, total choline intake increased from 23.5 to 177 and then to 326 g/d, but duodenal flow only increased from 1.2 to 1.3 and then to 2.5 g/d, respectively (Sharma and Erdman, 1988b). In the latter study, the apparent rumen degradability of unprotected dietary choline was much greater than reported in beef steers (Atkins et al., 1988), and ranged from 95.0 to 99.2 per cent.
Because of the extensive degradation of choline in the rumen, estimates of duodenal flow can barely match the needs of choline that is secreted in milk, less so the total requirements of choline for an adult lactating cow. Concentrations of choline in milk vary substantially, and it unsupplemented mid lactation cows it ranged from 72 to 86 mg/kg (Deuchler et al., 1998). In another experiment with transition cows, mean milk choline concentration in unsupplemented cows was 99.7 mg/kg (Pinotti et al., 2003). Although the choline requirements for dairy cows have not been established (NRC, 2001), Erdman (1992) suggested that duodenal flow of choline represents less than 30 per cent of the daily needs. For the modern high-producing dairy cows, it would be expected that unless de novo synthesis is present, cows would be in constant choline deficiency. Although it is unlikely that choline deficiency is present in modern dairy cows, it is possible that supplementation of choline might benefit metabolism and health given its role in phospholipid synthesis and lipid transport. Some (Sharma and Erdman, 1988a) have suggested that dairy cows might have requirements for choline beyond the need for methyl group donors and it cannot be overshadowed by supplemental methionine.
Sharma and Erdman (1989b) in a series of experiments with ruminally fistulated dairy cows demonstrated that abomasal infusion of varying amounts of choline consistently increased yields of milk and milk fat, whereas dietary supplemented choline had no effect on production responses. Therefore, because choline either present in or added to the diet is degraded extensively in the rumen (Atkins et al., 1988; Sharma and Erdman, 1988b), it must be protected from degradation to be of value to dairy cows.
Effects on intake
Supplementing transition cows with rumen-protected choline before calving did not influence DM intake prepartum, but when supplemented after calving, it tended to increase postpartum intake (Lima et al., 2007). Feed intake responses to supplemental rumen-protected choline have been variable and some studies have reported no effects (Erdman and Sharma, 1991; Hartwell et al., 2000; Piepenbrink and Overton, 2003; Zahra et al., 2006), whereas others observed an increase in DM intake (Oelrichs et al., 2004; Chung et al., 2005). It is unknown the mechanism by which choline might influence DM intake, but it is plausible to speculate an indirect effect mediated by improved postparturient health. Differences in response to rumen-protected choline might also be related to the quality of the product used and method of protection against rumen degradation as differences in the degree of rumen-protection have been shown for different products (Kung et al., 2003). Nevertheless, if choline has a direct impact on the risk of ketosis and fatty liver, then it is expected that appetite might be improved in supplemented cows.
Milk production and composition
Increasing the intestinal supply of choline has usually improved milk production in lactating dairy cows approximately 7 per cent over controls (Baldi and Pinotti, 2006), and this improvement seemed independent on the dose of rumen-protected choline supplemented to cows, which ranged from 6 to 60 g/d.
Erdman and Sharma (1991) conducted 2 experiments to evaluate the effect of different levels of rumen-protected choline and interaction between choline and dietary CP level in cows past 5 wk of lactation. In experiment 1, milk yield tended to increase with feeding of rumen-protected choline, and intake of choline chloride ranged from 17 to 50 g/d. In experiment 2, increasing intake of rumen-protected choline from 0 to 55 g/d of choline chloride resulted in a linear increase in milk yield despite dietary protein content (Erdman and Sharma, 1991). Generally, yields of milk and 3.5 per cent fat-corrected milk in response to rumen-protected choline have either been unaltered (Hartwell et al., 2000; Janovick Guretzky et al., 2006; Davidson et al., 2008) or increased (Erdman and Sharma, 1991; Piepenbrink and Overton, 2003; Zahra et al., 2006; Lima et al., 2007; Davidson et al., 2008). Increased responses to rumen-protected choline have been observed in cows with body condition > 4.00 prepartum (Zahra et al., 2006); nevertheless, increases in yields of milk, 3.5 per cent fat-corrected milk and energy-corrected milk have also been observed regardless of body condition of cows at calving (Lima et al., 2007).
Increased yields of milk components from feeding rumen-protected choline have generally been the result of increased milk yield, with some effects on milk fat content, but little or no effect on milk protein content. Because choline is used for phospholipid synthesis, it has been suggested that supplementation with choline may facilitate lipid absorption and transport, thereby favoring milk fat synthesis (Erdman et al., 1984); however, more detailed work on blood lipids revealed that supplementation with rumenprotected choline did not alter the concentrations of different lipid fractions in plasma of lactating Holstein cows (Janovick Guretzky et al., 2006). In early lactation, plasma nonesterified fatty acids supplies the majority of fatty acids secreted by the mammary gland in dairy cows, and this can increase if weight loss postpartum is exacerbated. Because rumen-protected choline might reduce fat mobilization based on plasma nonesterified fatty acid concentrations (Pinotti et al., 2002; Cooke et al., 2008), it is possible that potential effects of choline on intestinal lipid absorption, hepatic triacylglycerol secretion, and subsequent transport to the mammary gland might be masked by the reduced availability of nonesterified fatty acids for uptake by the mammary cells.
Responses to rumen-protected choline might also vary according to the supply of methyl donors such as methionine or other cofactors associated with methylation such as folate and other B vitamins. Inhibition of endogenous choline synthesis from methionine in lactating dairy cows reduced yields of milk and fat-corrected milk when cows received methionine infusion, but not when they received choline (Sharma and Erdman, 1988a), confirming that methionine is an important methyl donor in lactating dairy cows. Nevertheless, multiparous cows fed diets formulated to be inadequate inmethionine had increased milk production when supplemented with rumen-protected choline, but not when supplemented with rumen-protected methionine or betaine (Davidson et al., 2008). The same response to rumen-protected choline was not observed in primiparous cows, likely because milk production was low, approximately 27.5 kg/d. Collectively these data indicate that improvements in lactation performance in response to choline supplementation may be attributed to a methyl donor sparing effect, but also a direct effect of choline on lactation. It is also possible that increases in yields of milk and milk components cause by supplemental choline can be the result of improved postpartum health. Cows at greater risk for lipid-related disorders may benefit more from rumen-protected choline (Zahra et al., 2006), and cows of greater production potential (Davidson et al., 2008), although response to rumen-protected choline has been suggested to be independent on production potential of the cows (Pires and Grummer, 2008).
Pires and Grummer (2008) summarized 16 treatments comparisons in which rumen-protected choline was fed to dairy cows. Milk production in those studies ranged from 22 to 44 kg/d, and most of them supplemented rumen-protected choline at 15 g/d of choline. In 6 of the 16 comparisons, milk yield was significantly increased, and in none of them was milk production reduced in cows supplemented with choline. The authors observed that positive responses to rumen-protected choline were more often observed in cows producing between 30 and 35 kg/d.
Improving postpartum health with rumen-protected choline
Coordinated adaptation to lactation is critical to health and productivity of dairy cows. Lipid metabolism during the transition period plays a key role in this adaptive process. Excessive mobilization of adipose tissue to support milk and milk fat synthesis in early lactation results in increased incidence of subclinical and clinical ketosis and fatty liver, both of which compromise reproductive performance of dairy cows.
Recently, Lima et al. (2007) conducted two extensive experiments to evaluate the effect of rumen-protected choline fed during the transition period on incidence of health disorders, lactation and reproduction of dairy cows. In experiment 1, 362 Holstein cows were assigned to treatments at 253 days of gestation, and treatment diets were fed until 80 d of lactation. The diets were exactly the same, but cows receiving rumenprotected choline were dosed daily with 15 g of choline/day. In experiment 2, 573 Holstein primigravid cows were assigned to treatments at 256 days of gestation, and treatment diets were fed only during the prepartum period. The diets were exactly the same, but cows receiving rumen-protected choline were dosed daily with 15 g of choline/day.
In experiment 1, feeding rumen-protected choline before and after calving reduced the incidence of clinical and subclinical ketosis and the proportion of cows with ketonuria (Table 1).
Table 1. Effect of feeding rumen-protected choline (RPC) on ketosis in dairy cows
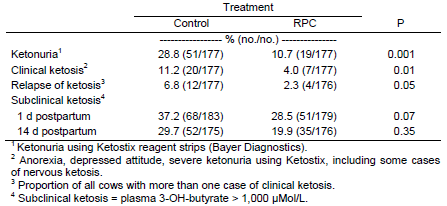
1 Ketonuria using Ketostix reagent strips (Bayer Diagnostics). 2 Anorexia, depressed attitude, severe ketonuria using Ketostix, including some cases of nervous ketosis. 3 Proportion of all cows with more than one case of clinical ketosis. 4 Subclinical ketosis = plasma 3-OH-butyrate > 1,000 μMol/L
In the same study (Lima et al., 2007) cows receiving rumen-protected choline throughout the transition period experienced lesser mastitis, although incidence of retained placenta, uterine diseases, and displacement of abomasum remained unaltered (Table 2). Because ketosis and mastitis were suppressed by feeding rumenprotected choline, the overall morbidity of cows in experiment 1 was reduced (Lima et al., 2007). In experiment 2, in which primiparous cows were fed rumen-protected choline only prepartum, incidence of ketosis and other postparturient diseases were similar between treatments.
Table 2. Effect of feeding rumen-protected choline (RPC) on health of dairy cows
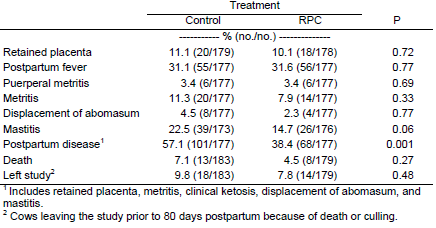
1 Includes retained placenta, metritis, clinical ketosis, displacement of abomasum, and mastitis. 2 Cows leaving the study prior to 80 days postpartum because of death or culling.
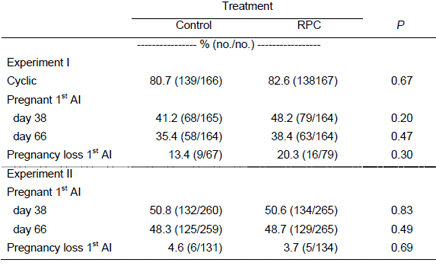
Reproductive performance of cows was generally not influenced by feeding choline. In experiment 1 (Lima et al., 2007), feeding rumen-protected choline did not affect the proportion of cyclic cows, proportion pregnant to the first insemination and pregnancy maintenance in both experiments (Table 3). These data suggest that feeding rumen-protected choline throughout the transition period improved postparturient health and minimized the risk for ketosis in dairy cows, but these benefits in health did not reflect improved reproduction. Furthermore, when fed only prepartum to primigravid cows, rumen-protected choline did not improve postpartum health and reproduction.
Table 3. Effect of feeding rumen-protected choline (RPC) on reproduction of dairy cows
Alleviation of fatty liver
Choline is used for phosphatidylcholine synthesis, a major phospholipid required for cell maintenance and replication. Phosphatidylcholine can also be synthesized from trans-methylation of phosphatidyletanolamine by methyl donors such as methionine (Vance and Vance, 2008). In monogastric animals, choline deficiency has been shown to result in hepatic lipidosis (Best and Huntsman, 1932). Choline can work as a methyl donor and spare methionine when amino acid supply is inadequate. Because choline is an integral component of the phospholipid phosphatidylcholine, it becomes a critical component for hepatic lipid metabolism. Phosphatidylcholine is needed for synthesis of very low density lipoprotein (VLDL), the lipoprotein responsible for export of triacylglycerol from hepatocytes.
In early lactation, dairy cows experience excessive mobilization of body lipid stores to support synthesis of milk and milk fat. This poses a risk for fat accumulation in the liver because the rate of fatty acid uptake by hepatocytes overcomes oxidation in the mitochondria or peroxisomes and export as VLDL. Excessive accumulation of fat in the liver has been associated with changes in blood biochemical parameters and reduced performance. Because choline might influence VLDL synthesis, it is possible that supplementation might minimize the risk of fatty liver.
Cooke et al. (2007) conducted two experiments to determine the effects of rumen-protected choline on triacylglycerol accumulation and depletion from the liver of dairy cows. In experiment 1, 24 dairy cows between 45 and 60 days prepartum were subjected to a feed restriction protocol and fed or not 15 g/day of rumen-protected choline. In experiment 2, 28 dairy cows between 45 and 60 days prepartum were subjected to a feed restriction protocol and fed or not 15 g/day of rumen-protected choline. In experiment 1, cows fed rumen-protected choline had hepatic tissue with reduced concentration of triacylglycerol during feed restriction and a period of negative energy balance. In experiment 2, cows fed rumen-protected choline experienced a greater depletion of accumulated hepatic triacylglycerol after feed restriction was suspended and cows were allowed to consume feed ad libitum. The data from Cooke et al. (2007) implied that rumen-protected choline can prevent and possibly alleviate fatty liver because of an increase rate of triacylglycerol depletion from the liver, at least when induced by feed restriction.
Yet, when fed to transition cows, rumen-protected choline has not reduced fat infiltration in the liver (Piepenbrink and Overton, 2003; Zahra et al., 2006), although it has increased glycogen content (Piepenbrink and Overton, 2003; Zahra et al., 2006). Interpretation of the findings by Piepenbrink and Overton (2003) suggest that feeding rumen-protected choline to transition cows increased the rate of VLDL secretion by the liver, which reinforces the in vivo findings by Cooke et al. (2007).
A problem with interpretation of average concentration of fat in the hepatic tissue is that liver has a wide variability in triacylglycerol content in the early postpartum period and cow with excessive concentration might mask a potential treatment effect. Furthermore, the high variability in triacylglycerol in the liver requires that many cows should be sampled per treatment to have a chance to detect differences.
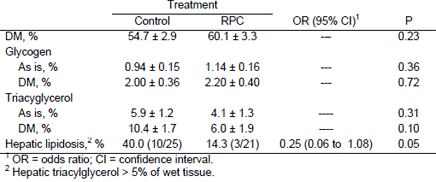
Lima et al. (2007) fed 15 g/d of rumen-protected choline to dairy cows starting at 25 d prepartum until 80 d postpartum. Liver tissue was collected between 7 and 10 d postpartum and analyzed for its chemical composition (Table 4). Feeding 15 g/d of rumen-protected choline did not influence liver DM and glycogen concentrations, but it tended to reduce the concentration of triacylglycerol in the hepatic tissue and reduced the risk of fatty liver.
Table 4. Effect of feeding rumen-protected choline (RPC) on hepatic composition in dairy cows
Summary
Although choline is not considered a required nutrient in the diet of dairy cows, duodenal infusion studies have shown that supplying choline to the small intestine for absorption increases milk yield and concentration of fat in milk. High producing dairy cows secrete 3 to 5 g/d of choline in milk, whereas duodenal supply is usually less than that. When fed in a rumen-protected form, choline improved milk yield in 38 per cent of the treatment comparisons from published literature.
Improvements in milk yield averaged 6 per cent, or approximately 1.5 kg/d. Feeding rumen-protected choline has been shown to decrease plasma concentrations of nonesterified fatty acids in transition dairy cows, and in cows subjected to feed-restriction. The reduced plasma concentrations of nonesterified fatty acids coupled with the improved hepatic export of triacylglycerol resulted in less hepatic fat concentration and reduced risk of fatty liver in some studies. Field studies with large number of cows have demonstrated that rumen-protected choline, when fed pre- and postpartum reduced the risk of clinical and subclinical ketosis and overall morbidity, but had no effects on reproductive performance.
Further Reading
| - | You can view a list of the full IFAS 20th Symposium proceedings by clicking here. |
May 2009

